


Nympheas, Claude Monet, 1922

Il faut donc repartir de la perception subjective des couleurs pour définir le vocabulaire qui permet de les décrire, via des expériences empiriques, plutôt que de la description physique de la lumière. Si Newton a posé les bases de la description des couleurs avec son cercle chromatique, qui montre qu’on peut « refermer » le spectre des couleurs de l’arc-en-ciel en un dégradé circulaire d’intensité constante passant par le violet, c’est un travail qui s’est surtout développé tout au long des 19e et 20e siècle. Dès 1802, Thomas Young soupçonnait l’existence de trois types de photorécepteurs dans l’œil humain, ce que des expériences de James Clerk Maxwell1 tendent à confirmer entre 1855 et 1872. Maxwell étudie la perception des couleurs et montre qu’on peut, avec seulement trois lumières de couleurs bien choisies, dans les bonnes proportions, produire une lumière dans une gamme très étendue de couleurs. Il utilise pour cela des instruments aussi simples et pratiques qu’ingénieux, par exemple une toupie colorée pour observer le mélange de trois couleurs primaires. Hermann von Helmholtz2 continuera d’explorer cette trichromie, tandis que dans le même temps, Ewald Hering3 développe une autre théorie des couleurs qui s’y oppose, non pas en partant de la composition de la lumière physique, mais plutôt de la psychologie et du langage. Il constate que le champ sémantique des couleurs, le champ chromatique, permet de décrire certaines transitions progressives d’une teinte à l’autre, par exemple allant du rouge au jaune, ou encore en allant du vert au bleu. Je peux tout à fait vous demander d’imaginer un bleu verdâtre ou un orange citronné. D’ailleurs, vous pouvez tout à fait concevoir qu’une orange mûrissant passe progressivement du jaune à l’orange, voire s’approche du rouge. Mais qu’imaginez-vous si je vous demande maintenant d’imaginer un rouge verdâtre ou bien une étape intermédiaire de la transition du vert au rouge d’une pomme qui mûrit ? Hering remarque ainsi qu’il semble impossible de concevoir certains mélanges de couleurs, en particulier du rouge tirant sur le vert ou du bleu jaunâtre. Il parvient ainsi à un système de description des couleurs sur trois échelles : une échelle d’intensité, de luminosité, allant du noir au blanc ; une échelle allant du bleu au jaune ; et enfin, une échelle allant du vert au rouge. Choisissez une couleur, n’importe quelle couleur : vous constaterez qu’il est possible de la décrire précisément grâce à ces trois échelles. Par exemple, la couleur de la table sur laquelle j’écris ces lignes, se situe plutôt en bas de l’échelle de luminosité, tirant très légèrement vers le jaune, et un peu plus franchement vers le rouge. Une couleur plutôt sombre, très légèrement jaune, et plutôt rouge, vous devriez avoir en tête le marron orangé du bois de cette table. Quoi qu’il en soit, il est beaucoup plus facile de décrire ou d’imaginer une couleur en utilisant ce système, qu’en donnant les trois quantités distinctes de rouge, vert et bleu de la trichromie. Si je vous avais dit que la même table est pour 80 % rouge, 20 % verte et 0 % bleue, je pense que vous auriez eu un peu plus de mal à imaginer du marron orangé. De la même manière, si je vous dis que la fleur que j’observe en ce moment est 50 % bleue, 40 % rouge, mais aussi 10 % verte, vous aurez sans doute plus de mal à vous figurer cette belle lavande que si je vous dis qu’elle est plutôt claire, lumineuse, et un mélange de bleu et rouge. C’est pourtant bien avec trois types de cônes et une sorte de trichromie que les yeux fonctionnent, alors pourquoi avons-nous tant de mal à décomposer les couleurs en trois signaux distincts correspondant à ces trois types de cônes ? Il y a en fait bel et bien un traitement supplémentaire dans les neurones du triple signal produit par les photorécepteurs, qui associe ces signaux en deux échelles bleu-jaune et verte-rouge, avec un traitement séparé de l’intensité, de la luminosité. C’est ce nouveau signal qui est transmis et traité par le cerveau, qui explique que nous voyons plus de contraste, plus de différence, entre le rouge et le vert, une paire « incompatible », qu’entre le vert et le jaune ou bleu. C’est au 20e siècle qu’Erwin Schrödinger4 a montré l’équivalence mathématique entre ce système de couleurs de Hering et la trichromie. On peut alors décrire l’arithmétique qui a lieu dans les neurones en deux équations simples : les trois signaux rouge (R), vert (V) et bleu (B), toujours positifs, sont combinés en trois autres signaux, trois échelles, rouge-vert (Rv), bleu-jaune (Bj), positives ou négatives, et luminosité (L), toujours positive, telle que Rv = R - V ; Bj = B - (R + V) ; et L = R + V + B. Ainsi, sur les deux premières échelles, une valeur positive indique respectivement une couleur plutôt rouge et plutôt bleue, tandis qu’une valeur négative donne une couleur plutôt verte et plutôt jaune, et la valeur zéro montre soit l’absence de signal, donc du noir, soit une couleur excitant égalitairement les trois types de cônes, qu’on percevra uniquement sur l’échelle de luminosité, allant du noir au blanc. On constate aussi aisément qu’une couleur composée seulement de rayons rouges et verts ne donne aucun signal sur la première échelle – Si R est proche de V, alors Rv = R - V ≈ 0. Ainsi, une couleur ne peut pas paraître un peu rouge et un peu verte à la fois. Le rouge-vert tout comme le bleu-jaune sont des couleurs dites « interdites » ; bien qu’on puisse physiquement concevoir un spectre lumineux qui correspondrait à ces couleurs, elles ne sont pas perceptibles et ne peuvent pas générer de qualia spécifique. Diverses expériences ont été tentées pour « faire voir » du rouge-vert ou du bleu-jaune à des humains, en superposant ces couleurs sur différentes parties de la rétine, ou en illuminant un œil avec chacune des couleurs par exemple, mais on n’arrive alors qu’à percevoir une superposition des deux teintes et non un nouveau qualia, une nouvelle couleur.


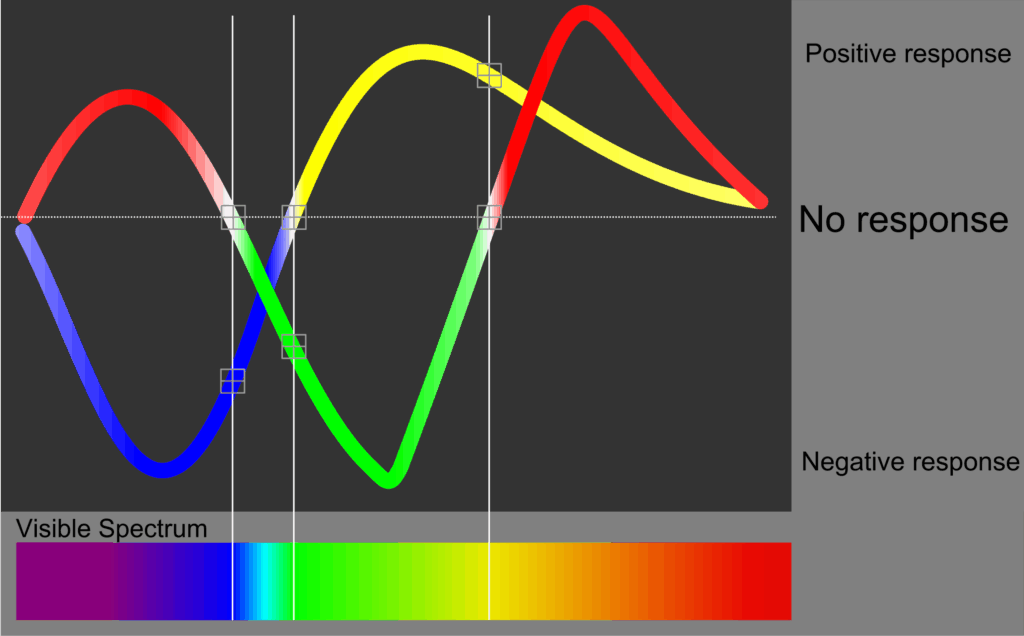
Ce système de perception des couleurs décrit par Hering au 19ᵉ siècle et qui peut s’expliquer par un traitement neuronal des signaux des photorécepteurs est à la base d’un système de couleurs conçu à la fin du 20ᵉ siècle qu’on appelle le NCS, pour « natural color system » (système de couleurs naturel), qui propose une représentation des couleurs différente de la trichromie additive Rouge-Vert-Bleu (RVB) des écrans numériques, et du système soustractif Cyan-Magenta-Jaune (CMJ) de l’impression numérique hérité de la peinture. Le système RVB reproduit les couleurs visibles en se basant sur le fonctionnement de la rétine et des cônes, en concevant un spectre lumineux calibré pour exciter spécifiquement ces cônes (et fonctionnant probablement mal pour les animaux qui ne discernent pas les mêmes sur nos écrans que dans la vie réelle). Le système CMJ s’attache lui à reproduire les teintes à partir de pigments prédéfinis et a nécessairement besoin d’un support aussi blanc et lumineux que possible ; on y ajoute souvent aussi un pigment noir pour augmenter le contraste qui peut être insuffisant en mélangeant simplement les trois couleurs primaires cyan, magenta et jaune. La correspondance, la conversion des couleurs décrites par le système de couleurs naturel (NCS) est relativement facile vers le système RVB mais plus complexe lorsqu’il s’agit de trouver la bonne proportion de pigments CMJ à partir d’une couleur décrite en termes d’échelle de luminosité, bleu-jaune et rouge-vert, ce qui fait que ce système NCS, plus intuitif, plus naturel comme son nom l’indique, et peu utilisé, n’ayant pas d’application pratique directe au-delà de sa capacité à décrire une couleur d’une manière plus directe pour les humains.
Le simple fait de capter et trier les longueurs d’ondes avec les cônes ne suffit donc pas à la vision des couleurs, qui est le résultat de cette combinaison des signaux dans les neurones. Le cas célèbre de l’artiste Jonathan I.5 le prouve : suite à un accident de la route sans gravité et ce qui a été diagnostiqué initialement comme une simple commotion, il a souffert d’une lésion cérébrale lui ayant fait perdre la vision des teintes, le plongeant dans un umwelt monochrome. Alors que ses yeux, ses rétines, continuaient de fonctionner de manière tout à fait normale, il n’était même plus capable d’imaginer les teintes, il avait totalement perdu ces qualia spécifiques. Il écrivit ces lignes, rapportées par Oliver Sacks et Robert Wasserman en mars 1986 :
J’ai vu des ophtalmologues qui ne savent rien de cette forme de daltonisme. J’ai vu des neurologues, sans succès. Sous hypnose je ne peux toujours pas distinguer les couleurs. J’ai subi toutes sortes de tests. Tout ce que vous voulez.
Mon chien marron est gris foncé. Le jus de tomate est noir. La TV couleur est un méli-mélo. Etc., etc.
Après 65 ans à voir les teintes, il était manifestement déprimé par son nouvel état, et il laissait transparaître à travers son discours posé une douleur, une peur, une tension intériorisées. En plus de la perte de vision des teintes, il a aussi souffert pendant quelques jours d’alexie, de cécité verbale, c’est à dire l’incapacité de lire ; bien qu’il voyait correctement les lettres latines, sont cerveau était soudainement incapable de les déchiffrer et il expliquait qu’elles lui paraissaient être du grec ou de l’hébreu. Il ne s’est par contre pas rendu compte immédiatement de son daltonisme, pourtant extrême ; le jour où il a décidé de retourner travailler, il a eu l’impression de conduire dans le brouillard bien qu’en ayant conscience du beau temps ce jour-là. Tout semblait brumeux, gris, indistinct. Il a été arrêté par la police en approchant de son studio, après avoir brûlé deux feux rouges, sans même s’en rendre compte. Entrant dans son studio, espérant que ce brouillard se dissipe, il a découvert ses peintures, connues pour leurs abstractions colorées, grises et vides de couleurs, incompréhensibles. « C’était horrible, dégoûtant », expliquera-t-il par la suite. Il ne pouvait plus supporter l’apparence des gens « comme des statues grises animées » ni son propre reflet, et n’était plus capable d’avoir des relations sexuelles, la couleur de la peau lui rappelant celle des rats. Et c’était toujours le cas les yeux fermés, toute son imagerie visuelle interne affectée aussi. La nourriture lui paraissait dégoûtante. Après une tentative, infructueuse, de peindre des fleurs colorées malgré son handicap, il s’est tourné vers la peinture en noir et blanc, et s’est réfugié dans son studio où il pouvait exercer une forme de contrôle, contrairement au monde extérieur qui l’angoissait, passant 15, voire 18 heures par jour à peindre, ses peintures montrant son évolution psychologique, depuis un monde brisé, angoissant, kaléidoscopique vers des représentations tout en mouvement de danseurs, de chevaux de course, pleines de vitalité et de sensualité alors que sa vie sociale et sexuelle revenait, avec l’atténuation de ses peurs et de sa dépression.


Des primates trichromates
Si ce système de la vision qui favorise la perception des contraste rouge-vert existe, c’est probablement qu’il a un intérêt évolutif pour l’espèce humaine.
Il semble que les premiers primates n’avaient que deux types de cônes6, les R et B. Il n’est pas possible de savoir précisément qu’elles étaient les couleurs qu’ils voyaient, mais la présence de seulement deux types de cônes réduit fortement la quantité de teintes pouvant être distinguées. Nous l’avons vu précédemment, le cône que nous appelons R chez l’être humain est sensible à une gamme de longueurs d’onde situées autour de 564 nm, allant en fait de ce que nous voyons comme vert jusqu’à l’orange, centré sur le jaune. Nous l’appelons R parce qu’il permet de distinguer le rouge, mais cette distinction du rouge ne se fait que grâce à la présence d’un cône V centré sur une longueur d’onde un peu plus faible, à 533 nm. Ainsi, si c’est uniquement le cône R qui est actif, mais pas le cône V, alors la couleur est d’une longueur d’onde plutôt longue, perçue comme rouge, tandis que si les deux types de cônes sont activés, la longueur d’onde est un peu plus courte et nous la voyons orangée ou jaune. Sans ce troisième type de cône, V, cette distinction est impossible, et, avec les cônes B, la vision chromatique se limite à deux teintes correspondant aux deux signaux des cônes, bleu-violacée et rouge, combinées à une échelle de luminosité. Ainsi, pour les premiers primates, mais aussi les chiens ou les chevaux, eux aussi dichromates, le ciel bleu contraste avec la prairie, celle-ci n’étant pas verte mais plutôt grise sombre ; il leur est difficile de discerner les fraises et framboises qui nous sautent aux yeux dans les sous-bois, et nos feux routiers n’auraient aucun sens pour eux. Il y a 29 à 43 millions d’années, au hasard des nombreuses mutations génétiques qui sont transmises lors de la reproduction, le gène codant l’opsine des cônes R chez une lignée de primates a muté, aboutissant à l’apparition d’un gène codant la nouvelle opsine, très semblable, des cônes V. Ces primates sont devenus trichromates et ont acquis la capacité de mieux distinguer les fruits qui contenaient moins de chlorophylle et étaient moins verts que les feuilles environnantes, favorisant ainsi légèrement leur capacité de survie, et transmettant donc ce nouveau gène aux générations suivantes. De la même manière, la reproduction des plantes aux fruits les plus oranges et rouges a à son tour été facilitée. On pense aussi que l’apparition de ce nouveau cône et de la trichromie chez les primates a pu les aider à mieux lire les émotions, signaux sexuels ou menaçants, sur la couleur de peau de leurs congénères, qui varie en fonction de l’afflux sanguin, et dont le rouge varie en fonction de la saturation en oxygène (c’est d’ailleurs en analysant cette couleur que nos machines médicales modernes surveillent cette saturation). Il semble d’ailleurs que la trichromatie chez les primates évolue de pair avec la disparition des poils sur le visage7.

Toutes les mutations génétiques ne sont pas favorables, et celles affectant les cônes peuvent provoquer une dyschromatopsie, qu’on appelle plus généralement le daltonisme, considéré comme une infirmité mineure. Dans des cas plus rares, le daltonisme peut aussi être la conséquence de lésions nerveuses ou oculaires. Il en existe différents types, dépendant surtout des types de cônes affectés, dont le plus courant est la deutéranomalie, où les cônes V sont déficients, mais pas complètement absents. Les personnes qui sont atteintes de deutéranomalie l’ignorent souvent, car elles conservent bien une vue trichromatique bien qu’un peu distordue. Les deux autres anomalies concernant les deux autres types de cônes existent aussi bien qu’elles soient plus rares ; ce sont la protanomalie, anomalie des cônes R, et la tritanomalie, celle des cônes B. Quand par contre les cônes V sont complètement absents, comme chez nos ancêtres primates, on parle de deutéranopie. C’est la forme dont était atteint John Dalton8, auteur de la première étude scientifique sur le sujet en 1798 et qui a donné son nom courant au daltonisme. Quand les cônes R et B sont absents, on parle respectivement de protanopie et de tritanopie. L’absence totale de vision des teintes, bien plus rare, est l’achromatopsie, qui peut survenir suite à une lésion cérébrale comme Jonathan I, ou être congénitale et est dans ce cas associée à une forte photophobie et une acuité visuelle très faible, mais aussi un nystagmus, des mouvements saccadés involontaires des yeux. Le daltonisme est plus répandu chez les hommes, parce que les gènes codant pour les cônes V et R sont situés sur le chromosome X ; les femmes en ayant deux exemplaires, il faut que les deux copies du gène soient défectueuses pour développer un daltonisme, alors que les hommes n’en possèdent qu’une copie unique.
Des couleurs au-delà du spectre visible

Prisonniers de notre umwelt, vivant dans notre monde propre et absolument incapables d’imaginer des couleurs autres que celles que nous percevons (l’histoire de Jonathan I. nous montre que même pour être imaginées, les couleurs doivent être traitées par les neurones en charge du traitement du signal visuel), nous pouvons simuler et comprendre, dans une certaine mesure, la vision de personnes atteintes de dyschromatopsie, ou éventuellement celle des animaux dichromates tels que chiens et chevaux, parce que ces visions reposent sur un sous-ensemble des signaux que les humains trichromates sont capables de percevoir. Nous avons tendance alors à oublier que le monde physique est bien plus vaste que ce que nous percevons, et qu’une infinité d’autres couleurs, de qualia, existe dans les consciences d’autres animaux. Nous pensons que la lumière solaire qui inonde la planète se décompose entièrement dans le spectre de l’arc-en-ciel. En 1912, Claude Monet9, alors en pleine création de sa série de 250 Nymphéas, est diagnostiqué d’une double cataracte, une opacification du cristallin. Devenu presque aveugle dans les années 1920, il se fait opérer des yeux, et se met à peindre un cœur bleu ciel aux nénuphars blancs. Il perçoit en fait une nouvelle gamme de longueurs d’onde dans la lumière, plus courte que l’extrémité bleu violacé de l’arc-en-ciel : l’ultraviolet. Cette perception reste toutefois extrêmement rare chez les humains. Mais si, au moyen d’un prisme de verre, vous décomposez la lumière solaire pour éclairer des fourmis, vous les verrez fuir cette zone invisible à vos yeux, l’extrémité ultraviolette du spectre. Les fourmis voient les UV qui inondent le monde10 et les océans, qui les dispersent facilement et en illuminent la vie sous-marine. Assez logiquement, les animaux percevant les UV sont nombreux, aussi bien dans les océans qu’à l’air. Les insectes sont trichromates, mais n’ont pas de cône sensible aux longueurs d’onde rouge : ils ont des cônes centrés sur les UV, le bleu et le jaune, et leur umwelt est bien différent du nôtre. Les fleurs, parsemant les prairies printanières de confettis multicolores, transformant les vallées alpines en splendides nuanciers répertoriant l’intégralité du spectre qui nous est visible, des bleus et blancs éblouissants de la neige et de la glace au sombre noir des rochers escarpés en passant par toutes les teintes de ces prairies, cachent aussi cette couleur supplémentaire à laquelle nous n’avons pas accès. Le tournesol, comme de nombreuses autres fleurs, et dont le jaune nous apparaît uniforme, est en réalité ultraviolet en son centre, et c’est cette couleur que les insectes butineurs repèrent pour savoir où est le nectar. Les insectes eux-mêmes sont souvent ultraviolets, ce qui fait bien l’affaire des oiseaux, eux aussi pourvus de photorecepteurs captant ces longueurs d’onde. De nombreux poissons, reptiles, les oiseaux et même certains mammifères voient aussi les ultraviolets, et les humains semblent bien être en fait une exception du monde animal. Nos cônes B sont aussi très légèrement sensibles à ces longueurs d’onde, mais elles sont en fait filtrées par les deux lentilles de nos yeux, la cornée et le cristallin. Si l’on perd ces lentilles comme Monet, après une lésion ou une opération, alors les cônes B peuvent percevoir les UV, qui se traduisent par un bleu ciel, laiteux ; mais ce n’est pas du tout comparable à la couleur perçue par les animaux dotés d’un type de cône spécifique à ces longueurs d’onde, produisant un quale qui nous est à jamais inaccessible et indescriptible.
Jusqu’en avril 2025, aucun être humain trichromate n’avait jamais perçu les qualia de couleurs qui correspondraient à l’activation d’un seul type de cône à la fois. En effet, tout rayon lumineux qui atteint la rétine est susceptible d’activer plusieurs cônes à la fois, même dans le cas d’un rayon ne comportant qu’une gamme très réduite de longueurs d’onde, puisque les différents types de cônes réagissent à des gammes larges et qui se chevauchent. Mais une expérience11 en avril de cette année-là, un dispositif ingénieux a permis de découvrir ces couleurs jusqu’alors impossibles. Au moyen d’un laser d’une longueur d’onde d’exactement 543 nm délivrée en micro-doses, cellule par cellule –
en liant le laser à la tête des participants et prenant en compte le mouvement des yeux – les chercheurs ont pu stimuler uniquement des populations de cônes V et étudier ce que les participants ont vu. Ils décrivent alors un bleu-vert saturé « au-delà du possible », autrement dit, un quale qui leur était jusque là inconnu. En situant cette saturation au maximum sur une échelle de 1 à 4, alors ils ont classé la plus saturée des couleurs naturelles à seulement 2,9 en comparaison ! C’est le seul exemple à ma connaissance d’une extension du champ des couleurs possibles pour un humain, lui faisant découvrir des couleurs au-delà de ce qu’il a toujours connu, mais cela ne semble pas pour autant introduire de nouvelles teintes, et ne nous permet pas d’imaginer, par exemple, ce que perçoivent les serpents à sonnette, dotés sur le museau de récepteurs infrarouges directement connectés à leur cortex visuel, les aidant à « voir » la chaleur de leurs proies12, ou ce que perçoivent les animaux dotés de cônes pour percevoir l’ultraviolet.
Tétrachromatie

Trichromates, les insectes perçoivent l’ultraviolet mais ne distinguent pas plus de nuances que les humains ; on peut encore simuler ce qu’ils voient, en remplaçant un quale par un autre, dans une sorte de correspondance entre les teintes que nous voyons dans les jaunes et rouges qu’ils ne perçoivent pas, et les ultraviolets qu’ils voient mais que nous ne percevons pas. Il est par contre tout à fait illusoire d’espérer se faire une idée du monde visuel des oiseaux, tétrachromates, qui possèdent des cônes aussi bien du côté ultraviolet du spectre que du côté rouge, en plus des verts et bleus. On considère que les monochromates discernent une centaine, ou en tout cas moins d’un millier, de nuances différentes, sur une unique échelle de luminosité, d’intensité. Les dichromates, avec une vision en deux dimensions, où chaque couleur doit être décrite au moyen de deux valeurs, deux échelles, peuvent discerner des dizaines de milliers de nuances, alors que les trichromates avec une troisième dimension en perçoivent des millions. À chaque nouvelle dimension, les nouvelles combinaisons possibles multiplient les perceptions, augmentant exponentiellement le nombre de nuances discernables, élargissant ce qu’on appelle le gamut, la gamme des teintes pouvant être perçues. Ainsi, nous discernons toutes les teintes de l’arc-en-ciel, mais nous sommes aussi capables de représenter toutes ces teintes sur un cercle fermé, reliant les deux extrémités rouges et bleues et formant le pourpre, une teinte qu’on qualifie alors de non spectrale. Le pourpre est en fait le résultat de la stimulation simultanée des cônes B et R, mais pas les V intermédiaires. Les tétrachromates multiplient encore le nombre de ces nuances, jusqu’à des milliards, multipliant la taille du gamut de leur umwelt, et peuvent théoriquement percevoir, en plus de la teinte ultraviolette, d’autres teintes non spectrales, combinaison des signaux des cônes qui ne se suivent pas, UV et vert, UV et rouge, et encore d’autres plus complexe impliquant trois cônes, UV, rouge et vert ; UV, rouge et bleu… L’ensemble des teintes perçues par les oiseaux ne serait pas représentable sur un cercle, mais sur un volume en trois dimensions dont notre cercle chromatique ne serait qu’un méridien, formant autant de teintes supplémentaires littéralement inimaginables. Si l’on peut simuler les visions aux gamuts plus réduits que la nôtre, monochromates, dichromates, et éventuellement dans une certaine mesure les visions trichromates différentes de la nôtre, le monde en tétrachromie de ma poule Agnès m’est définitivement inaccessible ; mais je comprends mieux maintenant son incroyable habilité à repérer quasi instantanément les plus petits insectes et araignées de mon jardin, aussi bien que les fraises qui poussent au bord de son enclos. Parce que la nature est pleine de surprises, quelques rares humaines sont aussi pourvues d’une vision tétrachromate. Il arrive en effet que les gènes codant les opsines présents sur les chromosomes X puissent différer sur l’une des deux copies. Une femme sur huit possède donc un quatrième type de cône. Il ne faut pas pour autant en déduire trop vite que les femmes ont une meilleure vision des couleurs que les hommes, puisqu’il ne suffit pas d’avoir ce type supplémentaire de cônes pour être tétrachromate ; il y a en effet beaucoup de conditions nécessaires à la vision effective de ces nouvelles couleurs. Ces types de cônes supplémentaires doivent être sensibles à des longueurs d’ondes suffisamment différentes des autres types, qu’ils soient correctement situés sur la rétine, mais il faut aussi que le système nerveux, le câblage de ces photorécepteurs, soit capable de prendre en compte ce nouveau signal et de le combiner aux signaux des autres cônes, ce qui fait de la tétrachromie humaine un phénomène rarissime, bien que documenté13. Mais les tétrachromates n’ont même pas conscience de leur différence ; vivant depuis toujours dans leur umwelt, elles n’ont aucune idée de l’effet que cela fait d’être trichromate, tout comme les trichromates n’ont qu’une idée très distante de l’effet que cela fait d’être dichromate, et ne vivent pas dans un monde qui leur parait comparativement incroyablement coloré. Si la tétrachromatie éveille votre curiosité, que pensez-vous de la douzaine de types de cônes différents de la mante de mer, elle-même arborant une splendide palette multicolore ? Il est en fait probable qu’elle n’ait aucune conscience de ces couleurs ; comme évoqué précédemment, il ne suffit pas de multiplier les types de cônes pour voir les couleurs, mais un traitement dans les neurones est nécessaire, et cette crevette ne semble pas posséder le système nerveux nécessaire à ce traitement et à la combinaison de tous ces signaux. Bien que possédant des yeux d’une complexité étonnante14, elle n’a pas un sens de la vision développé comme le nôtre, la complexité de ses yeux compensant en fait la simplicité de son cerveau, et chaque zone de ses yeux, chaque catégorie de photorécepteurs, remplit un rôle spécifique et spécialisé, automatique, sans nécessité de traitement complexe, ni conscient, et donc, sans couleur.
Et si la polarisation était une teinte ?
Nous découvrons que, d’une certaine manière, il existe un monde coloré bien plus vaste que ce que notre conscience nous met à disposition, comme si, enfermés dans nos esprits, nous ne percevions l’extérieur que par la petite lucarne de notre cellule. Tous les êtres vivants et voyants sur terre sont sensibles, globalement, à des longueurs d’onde allant de 300 nm (infrarouge) à 750 nm (ultraviolet), ce qui ne représente déjà qu’une petite partie du spectre de la lumière solaire. Mais les longueurs d’onde plus courtes et plus énergétiques, les rayons X et les rayons Gamma, sont filtrées par l’atmosphère. Quant aux longueurs d’ondes plus longues, les micro-ondes et les ondes radio, elles ne sont pas suffisamment énergétiques pour pouvoir correctement être absorbées par des opsines ; elles deviennent d’ailleurs trop grandes pour pouvoir être captées par des molécules microscopiques. Mais il existe tout de même une propriété des ondes lumineuses que nous n’avons pas abordée, mais qu’on pourrait tout à fait considérer comme une teinte supplémentaire. Prenez une longue corde et attachez là à un mur, puis secouez là dans tous les sens. Une onde, quelconque, se forme sur la corde, d’une manière similaire aux ondes lumineuses. Maintenant, secouez cette corde verticalement ; l’onde ne se produit plus que dans le plan vertical, et vous pouvez d’ailleurs faire tourner ce plan sous un angle quelconque, tout en gardant l’onde bien « plate ». Un phénomène comparable peut se produire avec la lumière, c’est ce qu’on appelle la polarisation15. Quand le plan ne tourne pas, qu’elle reste dans un angle précis, on dit qu’elle est linéaire, et si le plan tourne, elle est circulaire. La polarisation linéaire est très courante dans la nature, en particulier suite à sa réflexion sur l’eau ou des surfaces planes ou luisantes, comme le verre ou les feuilles de certaines plantes. Les insectes, les crustacés et les céphalopodes possèdent deux catégories de photorécepteurs sensibles à cette polarisation ; une catégorie détecte la polarisation sur un axe horizontal, et l’autre sur l’axe vertical. La combinaison de ces deux signaux permet d’en déduire l’angle, et tout porte à croire que les animaux la perçoivent comme une couleur supplémentaire, qui vient s’ajouter à celles formées par les combinaisons des différentes longueurs d’onde ! Ainsi, les pieuvres dichromates ajoutent en réalité une troisième dimension à leur vision, multipliant comme les trichromates les nuances qu’elles peuvent percevoir. Des dispositifs artificiels nous permettent assez facilement de détecter ou filtrer les lumières polarisées, comme des lunettes de soleil qui permettent en mer ou sur la neige de limiter l’éblouissement des réflexions, ou encore les lunettes permettant la vision en profondeur du cinéma en relief, 3D, ou les images destinées à l’œil gauche et droit sont projetées simultanément, mais polarisées verticalement ou horizontalement, et peuvent ainsi être filtrées différemment par chacun des deux verres des lunettes16. Mais là encore, il nous est impossible de distinguer cette potentielle couleur supplémentaire de l’umwelt d’autres animaux. La mante de mer, encore elle, est le seul animal connu à discerner la polarisation circulaire, où le plan de polarisation tourne, extrêmement rare dans la nature. Il se trouve que justement, une des seules sources de lumière polarisée circulairement dans la nature est la mante de mer elle-même, sur sa carapace ! Cette nouvelle couleur devient alors un moyen de communication discret et privé pour elles, utile pour la parade nuptiale et les rivalités entre mâles…
- James Clerk Maxwell (1831 – 1879) est un physicien et mathématicien écossais, principalement connu pouvoir avoir unifié en un seul ensemble d’équations l’électricité, le magnétisme et l’induction, pour former le modèle de l’électromagnétisme, en montrant par la suite, en 1861, que la lumière elle-même est aussi un phénomène électromagnétique. ↩︎
- Hermann von Helmholtz (1821 – 1894) est un physiologiste et physicien prussien ayant apporté d’importantes contributions à la dynamique des fluides, la perception du son et des couleurs et la thermodynamique. ↩︎
- Karl Ewald Konstantin Hering (1834 – 1918) est un physiologiste saxon, ayant principalement travaillé sur la vision des couleurs et la perception de l’espace. ↩︎
- Erwin Schrödinger (1887 – 1961) est un physicien et philosophe autrichien, prix Nobel de physique de 1933 avec Paul Dirac, pour l’équation d’évolution de la fonction d’onde des particules, à la base du formalisme théorique de la physique quantique. Il est aussi l’auteur de la célèbre expérience de pensée dite du chat de Schrödinger, développée à la suite d’une importante correspondance avec Albert Einstein en 1935. ↩︎
- Oliver Sacks et Robert Wasserman, The case of the colorblind painter, 1987. ↩︎
- Livia S. Carvalho et al., The genetic and evolutionary drives behind primate color vision, Frontiers in Ecology and Evolution, 2017. ↩︎
- Mark A. Changzi et al., Bare skin, blood and the evolution of primate colour vision, Evolutionary biology, 2006. ↩︎
- John Dalton (1766 – 1844) est un chimiste et physicien britannique, connu pour sa théorie atomique, premier modèle atomique ayant des bases scientifiques, publiée en 1808, ainsi que ses recherches sur le daltonisme. ↩︎
- Claude Monet (1840 – 1926) est un peintre français, un des fondateurs de l’impressionnisme, notamment connu pour ses séries à partir de 1890, où il peint le même sujet à différentes heures de la journée, à différentes saisons, peignant parfois des dizaines de toiles en parallèle. ↩︎
- Cette expérience a été faite dans les années 1880 par John Lubbock (1834 – 1913), préhistorien, naturaliste, mais aussi banquier et homme politique anglais. ↩︎
- James Fong et al., Novel Color via stimulation of individual photoreceptors at population scale, Science Advances, 2025. ↩︎
- Résultat de leur incandescence, surtout dans l’infrarouge pour les mammifères de quelques dizaines de degrés Celsius, comme décrite par le modèle du corps noir que nous avons abordé précédemment. ↩︎
- Gabriele Jordan et John Mollon, Tetrachromacy: the mysterious case of extra-ordinary color vision, Current Opinion in Behavorial Sciences, 2019. ↩︎
- Lire à ce propos le troisième chapitre de l’excellent livre d’Ed Yong, Un monde immense, comment les animaux perçoivent le monde. ↩︎
- Dans un modèle ondulatoire ; la polarisation est en fait liée au spin des photons vus comme des particules quantiques. ↩︎
- Au prix d’une division par deux de la luminosité perçue et du contraste, qui doit théoriquement être compensée par une projection plus puissante. ↩︎


Laisser un commentaire